|
2016年10月26日——美国ALS协会研究部门很高兴他们的研究投入发挥了功效。最近发表于顶级科学期刊《美国国家科学院院刊(PNAS)和《细胞(Cell)》的两项研究是由ALS协会支持的。第一篇论文由麻省大学医学院Zuoshang Xu博士和John Landers博士等研究人员发表,展示了一种表现出与人类疾病相似特征的新型profilin1(PFN1)小鼠模型,强调了开发新型动物模型对于理解ALS疾病过程和发现新疗法靶点的重要性。第二篇论文由圣裘德儿童研究医院J. Paul Taylor博士及其同事们发表。揭示出一种涉及C9ORF72扩张突变的新疾病机制,该机制将作为ALS疗法靶点。
9 z$ U4 a4 B9 I, o
新Profilin1小鼠模型复制ALS的多个方面 6 o! @9 c% |2 r9 C# o! Z
到目前为止,SOD1突变老鼠和一系列新兴ALS动物模型已经复制出人类ALS的主要特性:运动神经元变性退化、进行性无力和早逝。由于基因(遗传)突变只占全部ALS病例的一小部分,因此研究人员们一直在试图开发新的动物模型。在一项新的研究中,Yang等研究人员提出一种新的模型,也显示出这些特征,并让人联想到人类疾病的细胞特点。他们建立了突变ALS基因profilin1(PFN1)小鼠模型,这是一种罕见的ALS遗传原因——只有2%遗传性ALS患者携带该突变。 0 k$ V. t- L+ {3 a3 e4 c
! e8 j7 [) n' l2 o; r
John Landers博士和Zuoshang Xu博士
* Q* C. D7 Q9 q" w6 b
PFN1是一种肌动蛋白-结合蛋白,负责维持细胞的形状,对神经元轴突的维护和生长是重要的。他们表明,表达突变PFN1的小鼠会发展出与年龄有关的无力并最终瘫痪麻痹。他们也描述了脊髓中进行性变性退化的运动神经元的细胞特点。在症状出现后,运动神经元还发展出聚合物(蛋白质团块)。这些聚合物可能干扰运动神经元的蛋白质降解(回收)机器。
+ [2 n! Z5 x! v
还需要继续研究以确定PFN1诱导运动神经元毒性的确切原因。研究人员们计划使用这个模型进一步揭示ALS基础性的细胞通路。还将把这种新模型的进行性变性退化与SOD1小鼠模型进行比较,以理解这两种动物模型的相似与差别,从而为ALS疗法新靶点提供线索。
$ `4 j7 A& {1 e) T4 L2 Z" |; z
! U8 W( U1 k1 s7 u# O 揭示新的C9ORF72疾病机制
7 a/ @( i) ]8 { j. W
C9ORF72突变扩张是遗传性ALS和额颞叶痴呆(FTD)的最常见原因。这种突变涉及40%遗传性ALS和FTD病例以及5-10%散发性(没有家族史)ALS病例。C9ORF72基因的突变会导致“二肽重复蛋白质(DPRs)”的产生,之前的研究表明这些蛋白质对运动神经元具有毒性。 4 j1 m( f0 `% I% {0 m H
) ~0 v" y7 L2 x2 _9 [
+ ]: R) S* {# A; h3 z
Lee等研究人员通过探索DPRs相互作用的蛋白质类型深入研究这些蛋白质如何对运动神经元具有毒性。他们表明,两种都含有精氨酸的DPRs,会与多种RNA结合蛋白相互作用。这些RNA结合蛋白对处理和读取基因的遗传指令是重要的。特别地,他们发现DPRs与经常调节无膜细胞器功能的RNA结合蛋白相互作用。 ( k* c/ I/ `& i. t
无膜细胞器的例子包括核仁(负责制造构建蛋白质的核糖体)、应激颗粒(在细胞处于压力下时作为RNA临时存储地点的细胞内结构)和核孔复合体(分子在细胞核和细胞质之间穿梭的大门)。重要的是,DPR相互作用会扰乱这些无膜细胞器的动力学、物理性质和/或多个功能,潜在导致细胞毒性。 ) R; z$ v: e( ?% L) l& I# F
) O0 P# `( p1 U# l5 O) k9 u6 j3 y
这项研究为C9ORF72突变如何导致ALS提供了新的启示。研究结果还指向了另一个ALS疗法靶点——防止DPRs干扰RNA结合蛋白以停止对无膜细胞器的负面影响。这项研究还表明,无膜细胞器动力学和功能的破坏,可能存在共同通路,重叠在其他形式ALS中。使得这些研究结果对于整体理解疾病甚至更为重要。 $ f/ j: h F% B* I) X* D
) l% c3 D1 J7 I5 X( F' Z! @; T& Q/ B1 Y% t% p
0 P$ ^5 |$ m, @2 R | 




 发表于 2016-12-14 17:45:38
发表于 2016-12-14 17:45:38






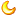
 发表于 2016-12-14 19:36:30
发表于 2016-12-14 19:36:30
 楼主
楼主

{kind=link}